Arthropoda (členovci) je největší a nejrozšířenější kmen bezobratlých živočichů. Je také velmi rozmanitý, co do velikosti (0,2 mm - 3m, např. fosilní hrotnatci) a bohatosti tvarů, že se s nimi nemůže rovnat žádná jiná skupina bezobratlých. Tělo členovců je vždy dvoustranně souměrné a výrazně článkované. Na tělních článcích jsou obvykle různé přívěsky, utvářeny podle funkcí, jež vykonávají (tykadla, čelisti, dýchací a pohybové orgány). Tělo je kryto pevným, často bílkovinným pancířem (chitinem), který bývá ježtě vyztužen tenkými vložkami uhličitanu vápenatého. Tento krunýř se při růstu musí svlékat. Tělo členovců je složeno ze třech základních částí - hlava, anterální hlavový štít (cephalon nebo caput), trup (thorax). Ten někdy splývá s hlavou v tzv. hlavohruď (cephalothorax - např. Ostrorep) a nakonec zadeček, posterální ocasní štít (abdomen - např. u mravence nebo pygidium - u trilobita). Nejprimitivnější členovci dýchají celým povrchem těla, u ostatních jsou vyvinuty různé typy dýchacích orgánů (žábry, vzdušnice). Pro poznání života v našich mořích starších prvohor jsou důležití představitelé tří podkmenů: Trilobitomorpha - trojlaločnatci, Chelicerata - klepítkatci a Crustacea - korýši [1].
Korýši (Crustacea) jsou velká skupina členovců (zpravidla klasifikovaná jako podkmen, přestože fylogenetickými analýzami byla odhalena její nepřirozenost) zahrnující asi 52 000 druhů. Patří k nim i velmi známé druhy živočichů, jako jsou krabi, humři či krevety, ale i některé velmi malé druhy, jako buchanka či perloočka. Korýši žijí především ve vodním prostředí, a to ve sladké či slané vodě, ale některé skupiny se adaptovaly i k životu na souši (svinka, stínka). Věda zabývající se korýši se nazývá karcinologie, případně malakostratologie či krustaceologie [2].
Klepítkatci (Chelicerata) jsou podkmen členovců. V současné době zahrnují jednu velkou třídu pavoukovci (arachnida - známe asi 73 000 druhů a patří mezi ně především suchozemští pavouci), jednu menší nohatky (Pycnogonida - známe zhruba 1000 druhů, jsou to mořští členovci s nápadně tenkým tělem, jejich velikost se pohybuje od 1-10 mm do 70 cm) a jednu téměř vymřelou hrotnatci (Merostomata - to je kdysi bohatá, ale dnes již téměř vymřelá třída mořských členovců, dnes zahrnuje 4 žijící druhy patřících do řádu ostrorepi (Xiphosura), přezdívaném také "živoucí fosílie", největší dosud žijící zástupci mohou měřit až 60 cm.
- primitivní formy klepítkatců, žijí v moři, zahrabáni v písku či bahně
- jsou to dravci, loví měkkýše a larvy bezobratlých
- tělo je kryto širokým plochým štítem - dorzoventrálně zploštělé, mají složené oči i jednoduché oči
-mají 1 pár končetin - klepítka a 5 párů kráčivých končetin (makadla nejsou rozlišena)
- zadeček nese končetiny s žábrami a dlouhý mečovitý výběžek
- po spáření vyplouvají samičky na pláže a zahrabávají vajíčka do písku (to je velmi zajímavé, protože mnozí trilobiti to mohli dělat podobně)
- vývoj je nepřímý, larvy, které připomínají trilobity, přelézají z pevniny do moře
- v současné době, díky znečištění moří mizí, přežívají jen 4 druhy- ostrorep americký (Limulus polyphemus), ostrorep východoasijský (Trachypleus tridentatus), ostrorep molucký (Carcinoscorpius rotundicauda) a ostrorep velký (Trachypleus gigas) - řadíme je mezi živé fosílie,( pozůstatky z devonského období -prvohory)
- velikost recentních druhů 60 - 90 cm
- Zástupci prvohorního řádu, tzv. kyjonožci, dorůstali délky přes 2 m. Jsou pravděpodobně suchozemskými pokračovateli vyhynulých trojlaločnatců a nemají společné předky s korýši a vzdušnicovci. Je jich známo asi 36 000 druhů. Je velmi pravděpodobné, že prvohorní trilobiti předali část svého genofondu právě těmto klepítkatcům.
Nás ale nejvíce zajímá první z těchto tří podkmenů, trojlaločnatci. Do tohoto podkmene patří dvě třídy, z nichž první, tzv. Trilobitoidea - trilobitovci, je dost nesourodá a do značné míry umělá. Sem patří členovci, jež s trilobity sice úzce souvisejí, ale přece jen se k nim nemohou přiřadit a patří spíše do tzv. problematik. Jsou to například některé fosilie z burgesských břidlic, jako Marella, jí příbuzná Furca (známá z českého ordoviku) nebo Duslia. Druhou třídou jsou konečně Trilobita - trilobiti [4].
Obecně můžeme trilobity charakterizovat jako starou, vymřelou třídu prvohorních členovců (arthropoda), která naši planetu obývala od kambria (jeho počátek je kladen před cca 542 mil. let) až po období permu, kdy na konci permu proběhlo velké masové vymírání (V důsledku kataklyzmatických změn vymřelo statisticky 77 % až 96 % všech vodních a suchozemských druhů! Vymřeli například poslední trilobiti, graptoliti, Pancířnatci a trnoploutvé akantódy. Obojživelníci subkategorie Lepospondyli a mnohé skupiny plazů. Vymírání postihlo i planktonické dírkonošce Fusulina). Konec permu je kladen 252 milionů let (Ma) zpět. Obývali tedy světová moře po dobu celého paleozoika (prvohor), což znamená, že zde byli nepřetržitě téměř 300 miliónů let, což je v porovnání s lidmi doba více než úctyhodná.
Jelikož jde o vyhynulé živočichy, jen stěží bychom dnes našli jejich blízké příbuzné. Přesto byl v 19. století v Bransfieldově průlivu v Antarktidě vyloven podivný přibližně 4 cm velký členovec, nápadně podobný trilobitům, a proto byl přírodovědcem Jamesem Eightsem nazván Serolis trilobitoides Eights, 1833 (obr. 1). Jde sice o také členovce, patří ale do podkmene Custacea - korýši. I když se také objevili v prvohorách, tvoří samostatnou vývojovou větev, jež na rozdíl od vyhynuvších trilobitů, přežili do současnosti. Trilobiti byli výhradně mořskými živočichy, což je zřejmé z nálezů, kde jsou spolu s typickou mořskou fanou: korály, ramenonožci, mlži a plži. Na druhé straně jsou doměnky, že některé druhy, především v kambriu a permu, se mohli adptovat sladkovodnímu způsobu života.


Obr. 1 - Serolis trilobitoides.
HISTORIE - stručná historie výzkumů a publikační činnosti týkající se českého barrandienu
Tělo trilobitů - jak již sám latinský název napovídá (tri - tři, lobos - lalok nebo tres lobus) se skládá ze tří částí, tzv. laloků. Ve vertikálním směru lze exoskeleton rozdělit na tři části. Mediálně (středem) probíhá axiální lalok a laterálně (postranně) dva pleurální laloky. Od tohoto rozdělení nesou trilobiti svůj název. Své jméno obdrželi trilobiti roku 1771 od paleontologa J. E. I. Walcha. Toto jméno se udrželo i přesto, že řada jiných autorů označovala trilobity jmény zcela odlišnými: trinuclei, entomostracites a Karl Linné je nazýval entomolithus. První publikovaná práce, která se zabývá staroprvohorní faunou Čech, je z roku 1770, pod komplikovaným názvem - Popisy vápencových lomů ležících u Prahy za vyšehradskou bránou s jejich zkamenělinami a jinými fosiliemi (přeloženo z německého názvu). Autorem byl jezuita a prof. matematiky na Pražské univerzitě Franciscus Zeno. Ten se zaměřil na fosilie z pražkého Podolí (Dvorce). Jeho vyobrazení jsou však natolik primitivní a názvy natolik smyšlené, že se dnes stěží můžeme domnívat, co vlastně popisoval. Pygidia trilobita Odontochile zveřejnil jako "Cacadu" a považoval je za měkýše s jednou miskou, nautiloidního hlavonožce popsal jako "Turbinites" atd. Tento stav tápání trval až do 20. let 19. století. Zlom nastává v roce 1822, kdy francouzský geolog A. Brongniart poprvé popisuje validní (platný) druh českého trilobita jako Asaphus Hausmanni, dnes známý jako Odontochile hausmanni (Brongniart, 1822). K dalším následovníkům, zkoumající českou staroprvohorní zvířenu (trilobity) patří např. Carl Zenker (ten se kromě trilobitů poprvé zajímá i o ostnokožce a popisuje např. lilijici, kterou nazve Scyphocrinites elegans, název i popis platí dodnes. Dále to je Kašpar hrabě Šternberk, německý badatel Ernst Beyrich, autorský tandem Ignaz Hawle a August Carl Joseph Corda až se na scéně konečně objevuje francouzský inženýr Joachim Barrande, jež vypracoval nejrozsáhlejší dílo v dějinách paleontologie, pojednání o českých fosiliích s názvem "Silurský systém středních Čech". Za jeho života vyšlo 22 svazků, ve kterých je popsáno a vyobrazeno celkem 3557 druhů. Dílo má přes 6000 stran textu a vysvětlivek k ilustracím a 1160 litografických tabulí s dokonalým, ač idealizovaným vyobrazením popisovaných druhů. Na každé tabuli je cca. 30 vyobrazení, na kterých se podílel sám Barrande (byl výborný kreslíř) [5].
ANATOMIE - pevné části těla
Ploché tělo bez orgánů, umožňujících rychlý pohyb, s očima umístěnýma na hřbetní straně krunýře - to vše nasvědčuje tomu, že trilobiti nebyli zdatní plavci. Mnozí byli i slepí či měli oči jen v juvenilních stadiích, nebo byl jejich hlavový štít přeměněn v lem, jímž rozhrabávali kal na mořském dně a hrabali v něm potravu, kterou byli především mikroskopické rostliny (řasy), drobní živočichové či odumřelé organické látky. Hřbetní krunýř, tvořený chitinem a zpevňovaný uhličitanem nebo fosforečnanem vápenatým (podobně jako u mnohých dnešních korýšů) byl většinou tenký: 0,05-0,2 výjimečně 0,5 mm silný, ale značně odolný a velmi dobře schopný fosilizace.
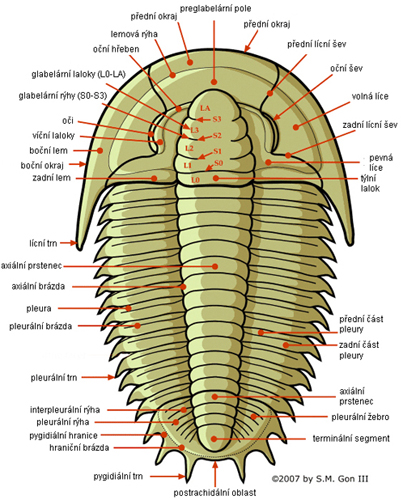

Obr. 2 - V horizontálním směru lze tělo rozdělit na anterální hlavový štít (cephalon), trup (thorax) a posterální ocasní štít (pygidium).
- Hlavový štít (cephalon) vznikl splynutím 5-7 článků, má většinou polokruhovitý či elipsovitý obrys a připojuje se zadním okrajem (tzv. týlním lalokem nebo prstencem) k prvnímu trupovému článku. Na hlavovém štítu lze rozlišit mediální část – glabelu – oddělenou od lící a ostatních částí hlavového štítu tzv. cirkumglabelární rýhou. Glabela můţe být členěna glabelárními rýhami do glabelárních laloků. Těsně ke glabele přiléhají pevné líce (fixigena) a společně s ní vytvářejí tzv. kranidium. Pevné líce (fixigena) obvykle nesou párové složené oči. Volné líce nazýváme librigena. Přední okraj cephalonu je vždy přehrnut dolů, takže vzniká zdvojení, tzv. duplikatura, na jejíž dolní okraj obvykle nasedá rozmanitě utvářený štítek - rostrum, na který navazoval hypostom, kryjící ústa.
- Trup (thorax) se skládá z různého počtu vzájemně kloubnatě propojených žeber - pleuronů. Obvykle jich bývá 8-16, ale počet článků se pohybuje od 2 (připustíme li, že agnostidi patří mezi trilobity) do více než sto (někteří trilobiti čeledi Emuellidae např. Balcoracania).

Obr. 3 - Balcoracania dailyi Pocock, 1970, zdroj: https://trilobites.naturalforum.net/t1321-famille-des-emuellidae-pocock-1970
- Ocasní štít (pygidium) bývá obvykle polokruhovitý či trojúhelníkovitý. Celý štít vznikl splynutím volně pohyblivých pleuronů. Počet segmentů ocasního štítu se pohyboval od jednoho až do třiceti. Stejně jako na trupu lze na ocasním štítu pozorovat axiální část a laterální pleurální části. Axiální část bývá často členěna na jednotlivé prstence, přičemž za posledním prstencem bývá ještě segment terminální. Okraj bývá buď rovný, či zdobený nejrůznějšími výběžky, případně i trny. U některých druhů vybíhá dozadu v menší či větší osten (tzv. kaudální trn). Na základě vzájemné velikosti hlavového a ocasního štítu bývá rozlišován exoskeleton mikropygální (ocasní štít výrazně menší než hlavový), subizopygální (hlavový štít nepatrně větší než ocasní), izopygální (hlavový i ocasní štít přibližně stejné velikosti) a makropygální (ocasní štít větší než hlavový) [6].
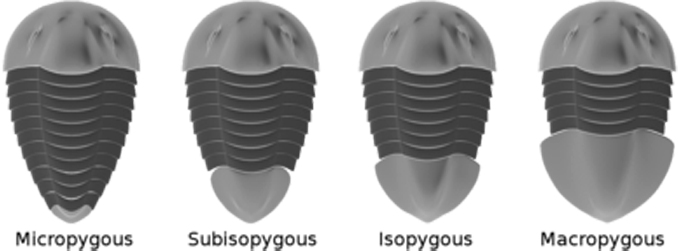
Obr. 4 - čtyři základní typy velikosi ocasního štítu.
ZRAK
Zrakové orgány trilobitů patří mezi nejstarší a vývojově nejzajímavější. Ne, že by se u těchto živočichů vyvinuli jako u prvních, ale poprvé zde došlo k tak velké tvarové a funkční diverzitě. Někteří trilobiti, většinou druhy žijící ve velkých hloubkách, byli zcela slepí. Jiní měli oči doslova „na stopkách “, tedy na zvláštních výrůstcích, které sloužili jako jakýsi periskop (Asaphus kowalewskii) . Díky nim se trilobit mohl ukrýt v kalu u dna a pozorovat dění ve vodě nad sebou. U některých druhů se oči vyvinuli tak, že mohli kolem sebe sledovat dění v úhlu 360º, aniž by se museli pohnout. Některé tvary očí mají tvar komolého kužele, podkovovitý, vejčitý nebo ledvinovitý. Některé jsou ploché, jiné naopak nápadně vyklenuté. Řada druhů měla drobné oči jen v mládí, ty později zakrněly, aby v dospělosti zcela zmizely. Oči se vyskytují na tzv. lícním švu, což je rozhraní, mezi pevnou a volnou lící. Oči trilobitů byly většinou složené z mnoha polygonálních čoček (podobné oči mají současní členovci). U trilobitů však byly dvojího, některé prameny uvádí oči trojího typu:
U tzv. holochroálního typu oko většinou pokrývala průhledná hladká blanka, která byla pokračováním tělní pokožky. Jednotlivá očka (omatidia) opatřená kruhovými nebo šestiúhelníkovými čočkami ležela těsně vedle sebe a mnohdy jich byl značný počet - v extrémních případech až 15 000, například u rodu Amphytrion, jehož nálezy známe i z Barrandienu – králodvorské souvrství. Naprostá většina trilobitů byla vybavena holochroálním typem očí.
Druhý typ očí, tzv. schizochroální typ, který se vyvinul, de fakto, pouze u fakopidních trilobitů, lze přirovnat k očím dnešních stonožek nebo štírů. Blanka překrývající oko, není u tohoto typu celistvá, ale má četné kruhovité nebo šestiboké otvůrky, ve kterých sedí čočky jednotlivých oček, oddělené sklerotizovanými stěnami. Celkový počet oček je pak výrazně menší, než u prvního typu, maximálně kolem 600. U těchto trilobitů můžeme pozorovat tendenci ke snižování počtu oček, tzv. redukci např. rody Acastoides, Struveaspis nebo Phacops (Procops).
Poslední typ, tzv. abathochroální oči, se našli pouze u agnostidních trilobitů, konkrétně u podřádu Eodiscina, trilobit Pagetia. Ti se vyskytovali v období kambria a ordoviku, pak definitivně mizí ze scény. Jelikož dnes agnostidi nejsou bráni jako trilobiti, je pchopytelné, že abathochrální oči nejsou mezi typy očí trilobitů řazeny. Jinak jsou to velmi drobná, oka, mají až 70 čoček. Ty se sami sebe nedotýkají a jsou odděleny tzv. interlensárním bělmem. Každá čočka má na svém povrchu vlastní rohovku. řazení omatidií působí poněkud chaotičtějším dojmem, než u předešlých dvou typů, ve skutečnosti jsou ale řazena ve směru horizontálních řad. Příslušnost agnostidů k trilobitům je ale v dnešní době sporná. Oči trilobitů byly jediné zrakové orgány v živočišné říši, jejichž čočky obsahovaly krystaly kalcitu. U některých taxonů došlo k sekundární ztrátě očí.
VELIKOST TRILOBITŮ
Průměrná velikost trilobitů sice kolísá mezi 2-10 cm, ale jsou známy i druhy měřící více než tři čtvrtě metru nebo naopak pár milimetrů. Mezi největší trilobity patřli někteří střednokambričtí paradoxidi, ordovičtí asafidi, lichidi, homalonotidi a devonští lichidi, homalonotidi a dalmanitidi. Někteří dosahovali velikosti kolem 70 cm. Největší popsaní trilobiti jsou v současnosti Isotelus rex Johnson a kol., 1988 – 72 cm, ordovik; Uralichas hispanicus Rabano, 1989 – 66 cm, ordovik; Terataspis grandis Reimann, 1943 – 50 cm, devon; Paradoxides davidis Bergstrom a Levi-Setti, 1978 – 37 cm, kambrium. Z českých jsou to Hydrocephalus carens Barrande, 1846 – 35 cm, kambrium; Birmanites ingens Barrande, 1846 – více než 30 cm, ordovik; Zlichovaspis (Z.) spinifera Barrande, 1846 – více než 30 cm, devon. Velmi rozmanité bylo utváření povrchu krunýřů. Jsou známy druhy zcela hladké či naopak poseté výběžky, z nichž původně pravděpodobně čněly smyslové orgány - brvy. Jiné druhy mají pleurony protažené v šavlovité různě dlouhé trny, které bezesporu sloužily coby obranný mechanismus při stáčení v případě napadení predátory. Trny vyzbrojení jedinci se tak nestávali snadnou kořistí velkých klepítkatců, ryb či dravých hlavonožců. Co se trnů týká, četl jsem nedávno tento výsledek studie: "Podle nedávné studie dvou britských vědců – biologů Roba Knella z londýnské Queen Mary University a Richarda Forteye z Natural History Museum vyplývá, že některé druhy trilobitů byly patrně nejstaršími účastníky soubojů v živočišné říši. Již před stovkami milionů let samci mezi sebou soupeřili o samičky. Vědci zkoumali různé druhy trilobitů, které se nacházejí v muzejních sbírkách, a zjistili, že některé z nich mají na hlavě podobné rohy jako dnešní brouci. Samci je v současnosti používají při bojích o nadvládu nad ostatními jedinci svého druhu a při zápasech o právo na páření. Vědci proto předpokládají, že rohy u trilobitů měly stejný význam. Přestože existuje celá řada dalších možných vysvětlení přítomnosti rohů u dávných živočichů, zdá se, že nejvíce pravděpodobný je právě jejich bojový účel. Výzkum se zaměřil na trilobity, kteří žili v ordoviku před 488 až 444 miliony let, ale není zcela vyloučené, že se závěry britských vědců mohou týkat i dalších druhů." zdroj: 21.století
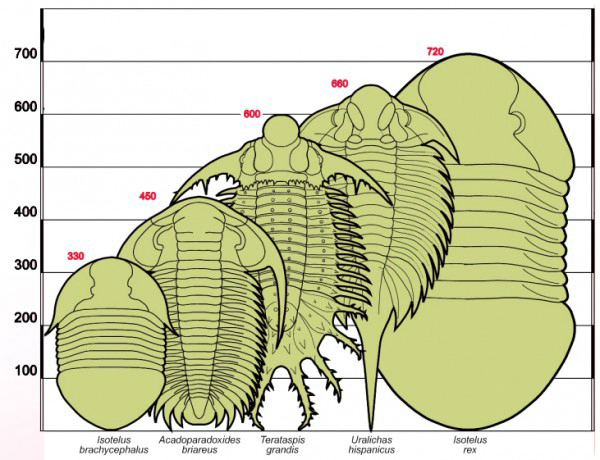
Obr. 5 - srovnání největších nalezených trilobitů.

Obr. 6 - Obří otisk trilobita Isotelus rex byl objeven na břehu Hudsonova zálivu, v blízkosti města Churchill, Manitoba, Kanada.

Obr. 7 - Trilobit Isotelus rex se ze skály musel získat po částech.
Isotelus rex je přiřazen k pozdnímu ordoviku Churchill River Group. Fosílie je lepená, neboť jak je zřejmé z doložených fotografií nálezu, nebylo možné získat otisk z břidlice v celku. To ovšem nijak nesnižuje význam nálezu. Při takto velkých otiscích je zcela běžnou praxí, je získávat postupně, po částech a následně je kompletovat.

Obr. 8 - Fotografie z náleru trilobita Isotelus rex.
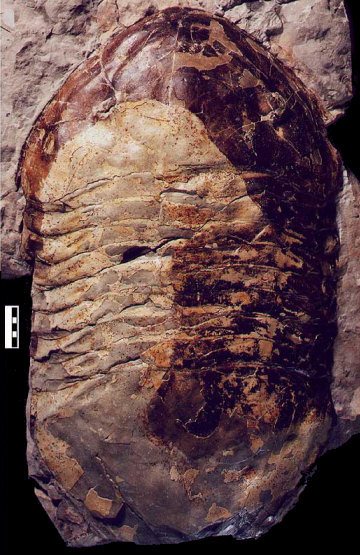

Obr. 9 - I když chybí kus pygidia, Isotelus rex je jednoznačně rekordmanem co se velikosti týče - délka 720 mm.
Ačkoliv se jedná o mimořádně velký nález trilobita, zachovali se na něm, i když to u velkých jedinců není zvykem, neuvěřitelné detaily, jako krásně zachovalé oči, velmi zřetelné volné líce atp.. Díky vysráženým minerálům má vcelku atypické a zajímavé žíhání.
Nález učinili Dave Rudkin (Royal Ontario Museum), Robert Elias (University of Manitoba), Graham Young (Manitoba Museum) a Edward Dobrzanske (Manitoba Museum) v roce 1999. Měří 720 mm na délku, 400 milimetrů na šířku (přes cephalon). Je součástí sbírek Muzea Manitoba ve Winnipegu (Kanada).
ANATOMIE - měkké části těla
O vnitřní stavbě jejich těla vypovídají jen nesmírně vzácné nálezy, které pocházejí např. z USA, Švédska či Austrálie (Emu Bay). Jedná se o tak jemnozrnné mořské sedimenty, že se nám přes propast věků dochovaly i ty nejjemnější části jejich těl: tykadla či končetiny, případně pocházejí z míst s tak unikátními fosilizačními procesy (lagerstätten), že se uvedené měkké části zachovaly. Běžné fosilizační podmínky neumožňovaly zachování měkkých částí těl trilobitů. Naprostá většina znalostí o těchto částech byly získány na základě analogie s dnešními členovci. Mezi tělní přívěšky trilobitů obvykle patří tzv. antény (tykadla - délkou se rovnaly nebo přesahovaly délku hlavového štítu a patrně byly vyuţžvány jako senzorické orgány), přívěšky plnící dýchací funkci a přívěšky motorické (končetiny). Doposud je známo kolem dvaceti taxonů trilobitů, u kterých byly objeveny tělní přívěsky [7]. Vynikající zachování vykazují nálezy ordovických trilobitů va státu New York v USA, především u olenidního trilobita [4]Triarthrus.

Obr. 10 - Zleva: Triarthrus eatoni se zachovalými měkkými částmi těla, New York - USA, ordovik, rekonstrukce trilobita Triarthrus becki. Zdroj: wikipedia.org
Znalosti o zažívacím traktu trilobitů pocházejí jednak z rentgenových snímků fosfatizovaných a pyritizovaných jedinců, ale také ze zachovalých výplní trávicí trubice u několika dalších taxonů. Ústní otvor trilobitů se nacházel na ventrální straně cefalonu pod hypostomem. Na něj navazoval krátký jícen směřující šikmo vzhůru a vpřed, kde byl připojen k žaludku umístěným pod glabelou (Chatterton et al, 1994). Žaludek se poté zužoval do střeva probíhajícího pod axiální částí trupu. To končilo patrně na ventrální straně ocasního štítu, kde mohl být anální otvor. Střevo bylo u některých jedinců v každém článku poněkud transverzálně rozšířeno (Gon, 2009). Toto rozšíření by mohlo odpovídat slepým výběžkům střev dnešních členovců, sloužící ke zvětšení trávícího povrchu [8]. U ptychopariidního trilobita Selenocoryphe platyura, který se nachází v Utahu, USA, se podařilo nalézt zbytky žaludku, trávícího traktu s postraními "slepými střevy" (obr. 11).

Obr. 11 - Selenocoryphe platyura, Millard County, Utah - USA, kambrium. Vlevo jsou zbytky zažívacího systému uměle obarveny modrou barvou.
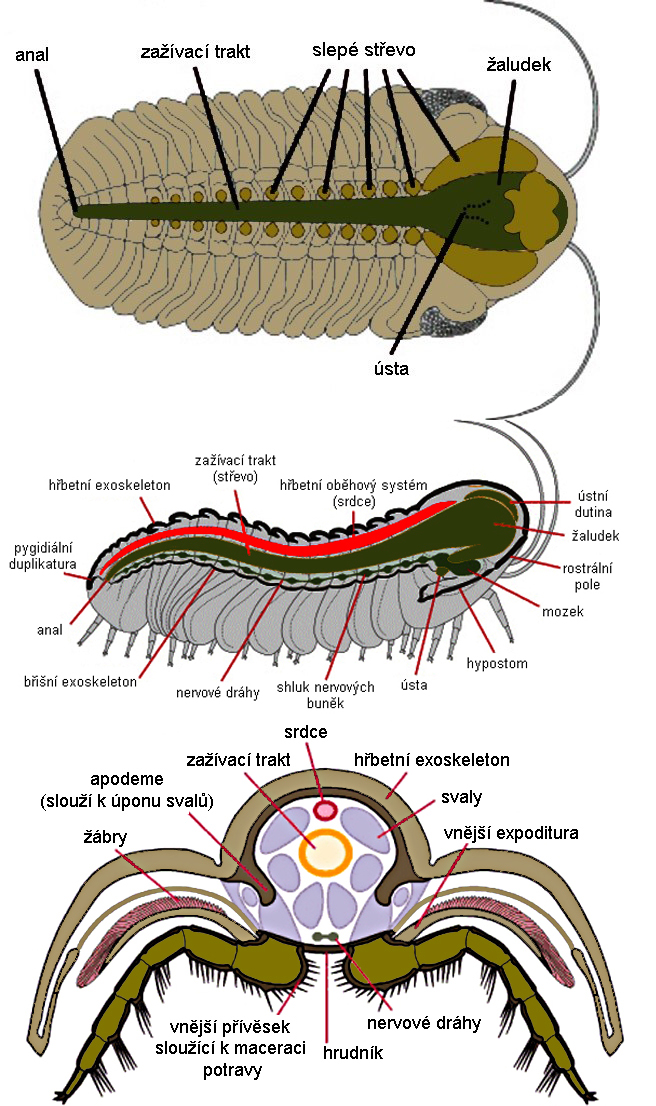
Obr. 12 - Rekonstrukce vnitřní stavby trilobita na taxonu fakopidního trilobita (Phacops). U tohoto druhu byl odhalen za pomocí rentgenu.
ONTOGENEZE - růst a vývoj
Trilobiti jsou vlastně jedni z prvních vymřelých skupin bezobratlých, u kterých byl popsán vývoj jedince od mikroskopické larvy, s pouze naznačenou tělesnou členitostí, až po dospělé jedince. Tělo se nejprve rozčlenilo na hlavovou a ocasní část. V další fázi proměny se tvořily trupové články. Jedním z nejznáměších příkladů, na kterých byl popsán vývoj trilobita, je práce Joachima Barranda, který jednotlivá stadia odhalil na trilobitu Sao hirsuta (obr. 13), z dnes již zaniklé legendární lokality "Pod hruškou" ve Skryjích. Barrande, i když nebyl první, byl určitě poprvé nejdůslednější. Mezi další rody, u kterých Barrande popsal ontogenetický vývoj patří Agnostus, Blainia, Paradoxides a Trinucleus.
Trilobiti byli pravděpodobně odděleného pohlaví, u některých druhů je popisován pohlavní dimorfismus. To znamená, že nejde jen o rozdíl v pohlaví jedinců, ale doprovázejí je i sekundární znaky, např. velikost trupu, tvarové odchylky (více v části Pohlavní dimorfizmus a rozmnožování), dále např. zbarvení - to samozřejmě dnes, díky stáří fosílií, již nelze dohledat. Onehdy jsem viděl dokument BBC, jak se jistý vědec pomocí nejmodernějších technologií, snažíl odhalit barvu peří podle nejlépe zachovalého Archeopterixe z německého Solnhofenu. Otisk tohoto "praptáka", jež je řazen již do třídy ptáků (Aves) - dnes velmi sporná a nedořešená otázka, někteří vědci jej pokládají spíše za dinosaura, nesl nejmenší detaily peří, co se prokreslení týče. Litografický kámen těžený v Solnhofenu má ale krásné krémové zabarvení do světlé, až střední neapolské žluti a toto zabarvení je defakto součástí všech místních otisků. Přeze všechnu snahu a dostupnou technologii, již měl vědec k dispozici a za kterou by se nemusely stydět ty nejlepší vědecké či lékařské kliniky, kýženého výsledku nedošel. Pigmentace peří se ztratila a soudobýmí dostupnými prostředky byla nedohledatelná. Ačkoliv Archeopterix lithographica žil na konci jury - období před asi 151-148 miliony let na území Německa, nepovedlo se. Je jasné, že podobné experimenty s mnohem staršími trilobity, jsou liché). Ale zpět k ontoganezi trilobitů. Z vajíček se líhly larvy. Počáteční larvální protaspidní stadium mělo krunýř rozdělený pouze na cephalon a pygidium. Krunýř v tomto stadiu dosahuje 0,25 – 1 mm. Meraspidní stadium začíná vznikem prvních trupových článků a končí dosažením plného počtu trupových článků. Dospělé holaspidní stadium mělo již ustálený počet článků, avšak i v tomto stadiu docházelo k růstu. Po něm následuje stadium gerontické, které se vyznačuje změnami vyvolanými stárnutím jedince.
V současné době se paleontologové snaží nahlížet na ontogenezi trilobitů z více úhlů. Například Hughes (2006) upozornil na to, že životní cyklus trilobitů lze rozčlenit nejen na základě artikulace (= vytváření volně pohyblivých článků, spojených kloubem), ale rovněž na základě segmentace (= přidávání nových segmentů) [9].
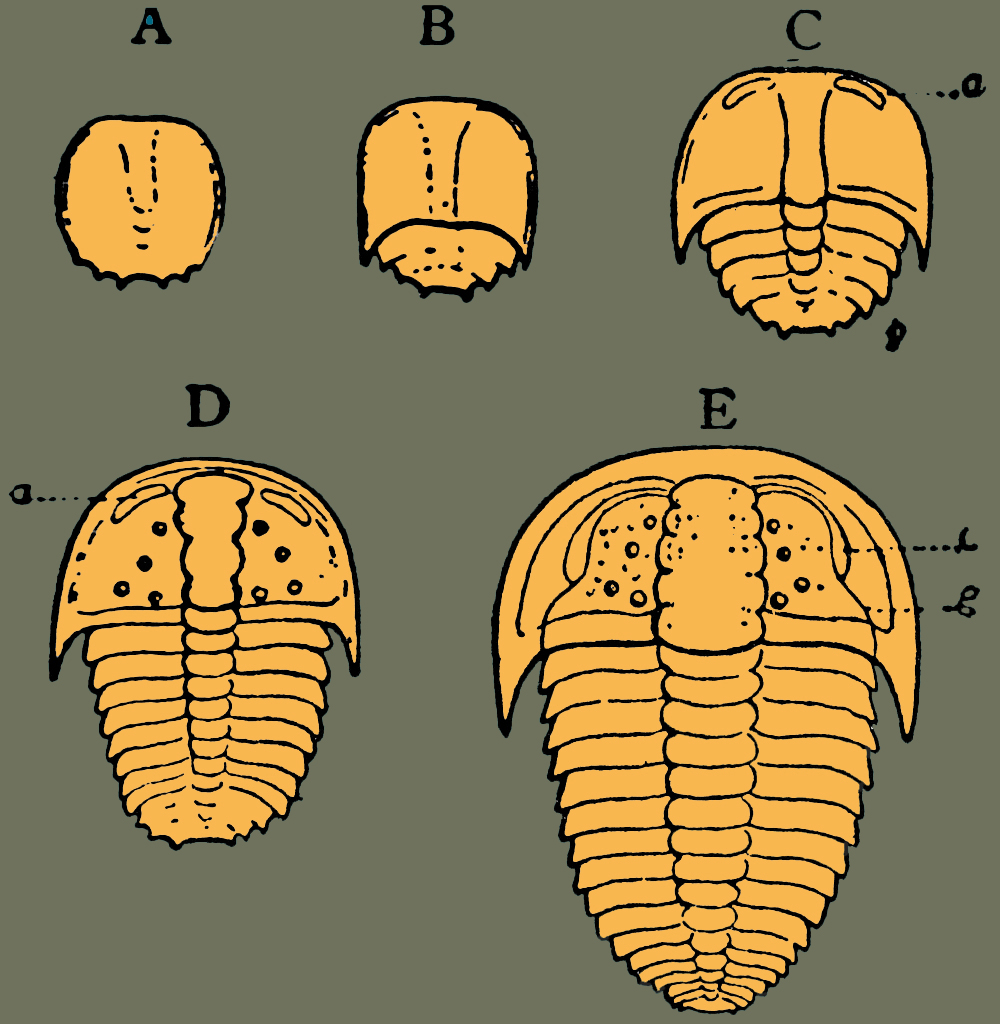
Obr. 13 - Ontogenetický vývoj rodu Sao. A – raně protaspidní stadium, B – pozdně protaspidní stadium, C – raně meraspidní stadium, D – pozdně meraspidní stadium, E – dospělý jedinec - holaspidní stadium (schematizováno, nejedná se o vědeckou ilustraci).
POHLAVNÍ DIMORFIZMUS A ROZMNOŽOVÁNÍ
O pohlavním dimorfizmu, potažmo rozmnožování trilobitů se ví velice málo. Stejně jako většina dnešních členovců i trilobiti se patrně rozmnožovali pohlavně (Chatterton a Speyer, 1997). Předpokládá se, že trilobiti byli odděleného pohlaví (gonochoristé). Pohlavní dimorfizmus byl u trilobitů popisován již Barrandem, který rozlišoval úzké (♂) a široké (♀) formy. Tyto rozdíly však byly později vysvětleny různou polohou fosílií vůči orientovanému tlaku v průběhu variské orogeneze (Šnajdr, 1990). Existuje ovšem několik dalších morfologických znaků, které různí autoři povaţují za pohlavní dimorfizmus (Chatterton a Speyer, 1997) [10]. Často je zmiňováno, že trilobiti jsou nejprimitivnější a nejstarší zástupci členovců. To, že patří mezi nejstarší členovce, je pravda jen částečná. Předchůdci členovců - trilobitů pocházejí, jak již víme, pravděpodobně z prekambria. Na první pohled se zdá, že mezi samcem a samicí nebyl žádný výrazný rozdíl. Je to proto, že se prozatím nenašly žádné průkazné konkrétní znaky této odlišnosti. Dle mého je ale občas potřeba číst mezi řádky. Říká se, že k pochopení současnosti musíme znát minulost. Totéž ovšem platí naopak. Abychom pochopili nebo se alespoň snažili pochopit to, co máme hledat, musíme nejprve studovat soudobé členovce a korýše a vlastně se to týká všech fosilních pozůstatků, jako jsou ostnokožci, hlavonožci, mlži, plži, ramenonožci, houby, korály a mnozí další živočichové. Na barevnou odlišnost pohlaví můžeme zapomenout, ta se po tak dlouhé době nedá dopátrat. Dozajista se totiž mnozí odlišovali právě na základě barevné různosti. Totéž můžeme pozorovat i u některých recentních krabů. Je faktem, že u soudobých členovců sice můžeme pozorovat tvarovou shodu mezi samci a samicemi, např. u právě zmíněných krabů, kde samce od samice rozlišíme buď podle zmíněného zabarvení, nebo tak, že pokud je otočíme bříškem nahoru, tak samice má ocásek ve tvaru širokého V, zatím co samec tvaru úzkého V. Zkuste ale takto otočit fosílii trilobita. Překvapivě ve většině prípadů narazíme na kámen. Měkká, spodní část trilobita se zachovala v tak málo exemplářích, že toto rozlišování neobstojí. Navíc nalezené měkké tkáně, respektive spodní strana trilobita, náležejí jiným rodům, mnohdy z jiné geologické epochy. To znamená, že je velmi málo pravděpodobné, že by se zachovaly dva takovéto otisky, náležející jednomu druhu a přitom bychom měli takové štěstí, že by se na jednom otisku zachovaly znaky samčí, na druhém samičí. Často v jednom souvrství nalézáme dva, velmi podobné druhy trilobita v různé velikosti, např. v Rejkovicích na Jinecku v zóně Eccaparadoxides. Ten rozdíl velikostí je ale markantní. Mám na mysli drobného trilobita Eccapardoxides pusillus a obřího, o to vzácnějšího Accadoparadoxida. Nemohlo by, při cílenějším studiu těchto "dvou druhů", jít zrovna o samce a samici? Předpokládám, že ten větší je samice a ti menší, kterých je, podstatně více jsou samci, kteří se snaží oplodnit vajíčka, pro přežití rodu a pak hromadně umírají, klesají ke dnu a fosilizují. Možná právě proto se nachází nepoměrně větší množství "samců" než "samiček - královen". Nepřipomíná vám tento scénář známou "telenovelu", jako vystřiženou ze soudobého světa hmyzu, například pavouků nebo včel? V mladší vrstvě jineckého a skryjsko-týřovického kambria se nalézají dva další adepti na to, abychom je při troše fantazie a odvahy "oddali". Jde o trilobity Conocoryphe a Ptychoparia. Tady zastávám podobný názor, a to ten, že ten evolučně vyvinutější, tedy Ptychoparia, který měl oči, by mohl být hledanou partnerkou, tak často nalézaných slepých samců Conocoryphe, kteří se "slepě" hnáni pohlavním pudem, snažili oplodnit nakladená vajíčka. Aby oplodněná vajíčka přeživší Ptychoparia mohla chránit, měla vývojovou devízu, oči. Tak bychom mohli samozřejmě pokračovat dál a dál, například v ordoviku celá řada Raphiophoridních, trinukleidních nebo akastidních trilobitů, o illaenidech ani nemluvě. Tam dle mého opět hraje roli různorodá velikost a četnost výskytu. Řada domnělých "párů" trilobitů je velká, mě ale trefili do oka dva, kteří se nalézají téměř vždy ve stejném souvrství, na stejných lokalitách. Myslím lichiidní trilobity Trochurus a Dicranopeltis. Jsou sice odlišní, ale zároveň velmi podobní, jako žena a muž. Prozatím jsem četl studii jen o tom, která pojednává zároveň o jejich příbuznosti (Lichiida), tak o jejich odlišnosti a v tom je možná kámen úrazu soudobé paleontologie. Každý sběratel fosílií se snaží, co to dá nalézt nový druh trilobita, trilobita, který se odlišuje velikostí, granulací, pygidiem, kranidiem, thoraxem, místem nálezu, počtem, tvarem a délkou ostnů na hlavě, trupu, zadečku. Málo se však už hledá podobnost, či příbuznost a tím nemyslím rodovou, ale spíše příbuznost druhovou.
Co se týká samotného rozmnožování trilobitů, odborníci věří, že samice trilobitů (my už také víme, že odborníci dnes netuší, jak přesně odlišit samice od samců) kladly vajíčka, která samci oplodňovali spermatem vypouštěním do vody. Byly nalezeny určité druhy s výrůstky na předním štítu, které by mohly svědčit o přítomnosti vaku, v němž samice buď nosila vajíčka, nebo své potomky, avšak ani tato domněnka nebyla potvrzena.
Obr. 14 - Ukázky trilobitů s chovným pouzdrem.
Obr. 15 - Kresba © 2000
S. M. Gon po Fortey & Hughes 1998
EKDYZE - svlékání kutikuky
Trilobiti, stejně jako ostatní členovci, svlékali během ontogeneze svůj exoskeleton v pravidelných intervalech. Růst a změny ve tvaru těla se odehrávaly během období mezi svléknutím a sekrecí následujícího exoskeletonu. Chování trilobitů během tohoto období není známé, protože měkký exoskeleton má menší potenciál k fosilizaci [11]. Vnější kostra neumožňovala plynulý růst. Celý proces byl řízen, podobně, jako u dnešních členovců. Základními hormony jsou svlékací hormon (ekdyzon) a juvenilní hormon (neotenin). Mají protikladné účinky. Juvenilní hormon udržuje znaky nedospělých stádií, blokuje vývin a přeměnu v dospělce. Ekdyzon vyvolává změny v pokožce a připravuje svlékání. Převládnutí účinků ekdyzonu – rozpuštění spodní části kutikuly, tvorba kutikuly nové, stará praská a je svlékána. Nová kutikula je nějaký čas měkká. Než ztvrdne chitinová vrstva, tzv. růst skokem.

Obr. 16 - Ukázka svlékání krunýře trilobita.
VOLVACE
Většina trilobitů měla schopnost, podobně jako někteří dnešní členovci, zaujmout víceméně kulovitý tvar a chránit si tak měkkou ventrální membránu a tělní přívěsky. Barrande (1852) rozeznává tři hlavní způsoby stáčení trilobitů: sféroidní, dvojité a terčové (diskoidní). Sféroidní stáčení bývá nejrozšířenějším typem. Posterální okraj pygidia se přitom dotýká anterálního okraje cefalonu. Distální konce pleur jsou těsně přitisknuty k sobě. Nejčastěji ho lze pozorovat u trilobitů s izopygálním (pygidium je stejně velké jako hlava) nebo makropygálním (pygidium je větší než hlava) exoskeletonem. Hughes (2003) uvádí obecný trend většiny trilobitů, kdy docházelo k postupnému zvyšování pygidiálních článků na úkor článků toraxu (tzv. kaudalizace), a tak mohlo být dosaženo izo- nebo až makropygálního exoskeletonu. Dvojité stáčení je popisováno u trilobitů s mikropygálním exoskeletonem (např. Ellipsocephalus). Pygidium se v tomto případě nedotýká okraje cefalonu, ale je spirálně stočeno pod něj. Při terčovém stáčení dochází k jednoduchému ohnutí toraxu a jeho přitisknutí k ventrální straně cefalonu. Pravděpodobně se jedná pouze o zvláštní případ sféroidního stáčení (Špinar, 1960). Ke zpevnění svinutí sloužila tzv. uzavírací brázda a uzavírací zářezy na antero-ventrálním aţ ventro-laterálním okraji cefalonu, do které zapadal posterální okraj pygidia (Henry, 1985).
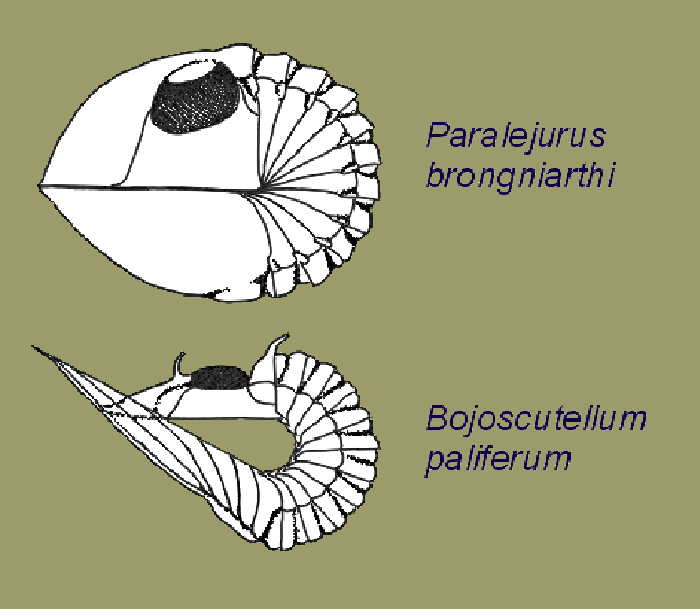
Obr. 17 - Ukázka volvace scutelidních trilobitů. Zdroj: https://www.kbi.zcu.cz/OB/veda/paleontologie/zoopaleontologie/trojlalocnatci/index.php
Obr. 18 - Neoasaphus Obr. 19 - Harpes
TAFONOMIE - vznik a způsob zachování fosilií
Trilobiti patří k nejhojnějším fosiliím staršího paleozoika, v malém měřítku mohou mít až horninotvorný význam. Nálezy celých exoskeletonů jsou však vzácné. Je to dáno malou odolností blanitých spojů mezi částmi exoskeletonu. Po uhynutí jedince blanité spoje podlehly rozkladu a proud nebo činnost jiných organismů vedla k oddálení osamostatněných částí krunýře. Při ekdyzi došlo již primárně k oddělení cephalonu a někdy volných lící od zbývajících částí krunýře. Běžně je pak nacházíme exuvie splavené na jedno místo a velikostně vytříděné. Původní barvy krunýře nejsou až na výjimky známé, ojedinělé nalézy ukazují na pruhy a kruhovité skvrny v řadách rovnobězných s průběhem pleur.
V písčitých sedimentech se exoskelety trilobitů často nezachovávají, o osídlení dna trilobitovou faunou však svědčí charakteristické stopy po pohybu (Cruziana, Rusophycus). Příčinou je chemické rozpouštěním kalcitických krunýřů v průběhu diageneze. Slabě sklerotizované končetiny a tykadla se zachovávají velmi vzácně. Průběh trávicí trubice byl zjištěn podle její odlišné výplně od okolního sedimentu, a to i z našeho území.
STRATIGRAFICKÝ VÝZNAM
Zkameněliny trilobitů svědčí nejen o druhové rozrůzněnosti a vývoji života jako takového, ale pomáhají také odhalit tajemství pohybu kontinentů. Je totiž známo, trilobiti žili na kontinentálních šelfech. Podle rozšíření několika typů fosilií lze vytvořit mapu oblastí, kde žili. Tyto mapy pak napomáhají ověřit i zdánlivě málo pravděpodobné teorie o kontinentálním driftu. Během dlouhé éry trilobitů prošel zemský povrch mnoha proměnami. Nálezy těchto vyhynulých členovců podpořily teorii o rozdílném geologickém původu Anglie a Skotska, které možná byly před 4 miliony let součástí dvou různých kontinentů, rozděleným oceánem lapetus. Tyto země se liší jak geologickou povahou oblastí, tak zkamenělinami trilobitů. Druhy, které se našly ve Skotsku, odpovídají druhům žijících v Kanadě a Grónsku (tyto oblasti kdysi tvořili Laurasii), zatímco v Anglii žili trilobiti v blízkosti kontinentu Avalonia. Několik set druhů má velký stratigrafický význam, zejména pro období kambria a ordoviku.
ÚSTUP
Existuje mnoho hypotéz, proč éra trilobitů na konci permu definitivně skončila. Žádná ale především uspokojivě nezdůvodňuje proč. První éra vymírání nastala na konci kambria. Jednalo se o tzv. první varilské vrásnění, kdy se posouvaly zemské desky. U nás jsou doklady pouze ze středního kambria (Jince, Skryje a pár méně známých lokalit). Z posledních nálezů z Maroka se ukazuje, že přechod ze svrchního kambria do ordoviku byl nakonec velmi pozvolný (souvrství Fezouta - Maroko). Dramatičtější to bylo na konci ordoviku. Proběhlo druhé varilské vrásnění a téměř celá populace živočichů zanikla. Silurské moře je v počátku známo jako nepříznivá oblast bez života. Podmořské sopečné plyny zahubily vše živé. Z této asi 60 metrové vrstvy jsou známi pouze graptoliti. Ostatní živočichové jakoby zmizeli a to na poměrně dlouhou etapu istorie. Ačkoliv Hans Zilmer by tuto etapu zkrátil, neboť vybuchne li sopka, je schopna během pár hodin zasypat vrstvu o 60 metrech, šlo o podmořské sopky, kdy se v této 60 metrové vrstvě objevil život v podobě graptolitů. Především to byla změna klimatu, doprovázena vysycháním moří a jejich těla, která nedávala velké možnosti vývoje, jako mnohem lépe přizpůsobivější plazi. Ačkoliv tito živočichové byli možná opravdovými vládci prvohorních oceánů, je málo pravděpodobné, že by se ještě dnes mohl ve světovém oceánu ukrývat jediný exemplář. Jedno je však zřejmé: pokud jde o schopnost přežít, patřili tito členovci k velmi úspěšným živočichům, nakonec se však ukázali úspěšnější organismy jiné, adaptabilnější na změny. Trilobiti se při svém třetím globálním vymírání nevzpamatovali a definitivně skončili jako překrásný doklad vývoje živých organismů.
"Trilobiti však na konci permu nevymřeli náhle, nýbrž tato skupina zaznamenávala výrazné úbytky na diverzitě (druhové rozrůzněnosti) ještě před touto biologickou krizí. Po slavné éře kambrické a ordovické je ústup trilobitů v siluru již patrný. Vzestup ve střední části spodního devonu, který započal po eventu hranice lochkov-prag (lochkov a prag jsou mezinárodně uznávané stupně spodního devonu), byl jen dočasný. V karbonu je již úbytek drastický. Globální event na hranici perm-trias neměl s trilobity příliš práce. Dokonce žádnou, neboť tato kdysi dominantní skupina živočichů nenápadně odešla ze scény ještě před tímto eventem.
Vezmeme-li v úvahu Český kras, je to jedna z nejdůležitějších oblastí na Zemi, kde lze zkoumat fenomén postupného úbytku trilobitů ke konci spodního a ve středním devonu. Tímto problémem se v poslední době zabývá jeden z nejvýznamnějších světových odborníků na devon, Prof. Ivo Chlupáč z Karlovy univerzity v Praze, jemuž tímto děkuji za cenné připomínky k textu. Jeho závěry, publikované v prestižním francouzském časopise Geobios (viz Chlupáč 1994), jsou více než přesvědčivé. Menší biologické krize (eventy vymírání) v devonu byly sice osudné pro čeledě trilobitů, ale ústup v podobě snižujícího se množství rodů a druhů byl zaznamenán rovněž v předstihu, tedy před těmito eventy. To je velmi důležité, protože nám to ukazuje, že ani tyto dílčí eventy nebyly příčinou všeobecného ústupu trilobitů, nejvýše jej určitým způsobem urychlily.
Spolu s úbytkem trilobitů šel ruku v ruce ještě jeden pozoruhodný fenomén, který rozhodně stojí za zmínku. Tak, jak se snižovala diverzita, zvyšovala se naopak tvarová rozrůzněnost (odborníci říkají, že byly vyhraněnější "morfotypy"). To je nesmírně důležitý a zajímavý postřeh. Nelze ho opomíjet, protože něco podobného lze pozorovat např. i u dinosaurů. Srovnáme-li životní osudy obou těchto notoricky známých skupin, seznáme, že jsou si podobné až zarážejícím způsobem: Trilobiti během prvohor postupně ubývají a jejich morfotypy se stávají stále výraznějšími. Konec jejich dlouhé éry přichází těsně před globálním eventem na hranici perm-trias, kdy se, zjednodušeně řečeno, dokončuje změna charakteru mělkých moří z původních rozsáhlých epikontinentálních ploch staropaleozoického typu v dnes dobře známé šelfy. Dinosauři sice výrazně neubývají, ale i tak lze pozorovat, že určité jejich skupiny do svrchní křídy nepřežívají. Morfotypy dinosaurů jsou v křídě zřetelně nejvyhraněnější. Konec jejich úspěšné éry učiní globální event na hranici křída-terciér, jehož sílu vyvolá (zdá se to být opravdu velmi přesvědčivé) pád velkého kosmického tělesa. Nejsou to ale jen trilobiti a dinosauři, kteří mají podobný trend ke stále vyhraněnějším morfotypům. Je to zákonitost univerzální. Všimli si toho už badatelé ke konci 19. století a slavný americký neolamarckista Edward Drinker Cope (1840-1897) nazval tento fenomén "zákon nespecializovaného předka" (Cope 1896). Spokojíme se s tím jako s podobností čistě náhodnou? Zamysleme se nad jinou možností. Letos vyšla zajímavá publikace v Knižním klubu: "Jak lidé umírají". Trpí sice módní nedbalostí mnohých našich současných redakcí (chybu zde naleznete rekordně takřka na každé stránce), ale je to jinak neobyčejně krásná a hodnotná kniha, která si zaslouží reklamu. Využil jsem zde parafrázi názvu této knihy a motto. Má to samozřejmě svůj důvod, neboť si uvědomuji, že je nutné se zamyslet nad tím, jak je možné, že dnešní věda nebere vůbec v úvahu možnost, že druhy stárnou - podobně jako jedinci. Kdo čte sci-fi literaturu, je dobře obeznámen s tím, že lidstvo je chápáno jako něco, co bude existovat navždy, pokud se ovšem nezničí samo či nezanikne zapříčiněním někoho jiného.
Vrátím-li se ke zmíněné Nulandově knize, zaujalo mne, že v současné medicíně existují v hrubých rysech dva směry myšlení, které se snaží vysvětlit proces stárnutí jedince. Jeden z nich, teorie opotřebování, zdůrazňuje neúnavné poškozování buněk ze dne na den až do okamžiku smrti. Druhý směr se přiklání k víře, že stárnutí je dáno geneticky, tj. předurčenou délkou života jednotlivce i jeho buněk a orgánů (do této druhé skupiny, velmi početné, se jasně hlásí i zastánci přírodního výběru a darwinovské medicíny - viz Nesse & Williams 1996). Zdá se být divné, že podobný stav nepanuje v současných teoriích o vymírání druhů. Analogie teorie opotřebování existuje v podobě neokatastrofismu a selekcionistické (darwinistické) teorie vymírání druhů neschopných konkurence, ale analogie teorie genetické zhola chybí. Vymizela (jakoby navždy) spolu s úmrtím německého paleontologa Otto Schindewolfa, který v padesátých letech tohoto století probojovával teorii stárnutí fylogenetických linií (tzv. typolýzu), a to v rámci své proslulé, dnes jednotně zavrhované, typostrofické teorie evoluce.
Každý slyšel o hromadných vymíráních druhů (mass extinctions), jejichž příčinou jsou katastrofy všeho druhu. Méně čtenářů ví o tzv. průběžných vymíráních v "pozadí" těch velkých (background extinctions). Je jich však nepoměrně více. Pokud je čtenář zvídavější, dávno se již informoval o tom, že i u těchto vymírání se obvykle uvažuje o nějakém zásahu zvenčí. Nejpozoruhodnější ale je, že mezi oběma typy není ve fosilním záznamu žádný ostrý předěl, ale plynulé kontinuum (Raup 1995).
I přes tuto fascinující skutečnost, kterou by bylo třeba rozebrat velmi podrobně, panuje stále kuriózní stav - dnešní věda považuje živočišné druhy za nesmrtelné. Jejich zánik může být způsoben jedině "špatným zacházením", a to ať už ze strany jiných druhů (David Raup poněkud nešťastně označuje tento způsob jako "špatné geny"), anebo ze strany nevlídného prostředí. Jinou možnost nebere v potaz, což je právě to, co může být pro laika nesrozumitelné. V takto načrtnutém scénáři pak David Raup ovšem dochází, na podkladě bohatých osobních znalostí statistiky, k závěru, že každý druh jednou vymře, neboť každá statistická "hra" (vskutku neobyčejně oblíbené slovo mezi současnými evolucionisty!) a pochopitelně i hra o život, dříve či později musí skončit fiaskem.
Nakonec se s tím lze smířit: jednak proto, že je obvyklé uznávat světové autority, jednak proto, že statistické vysvětlení má vskutku sugestivní logiku (každý přece ví, že v karetní hře nelze vyhrávat donekonečna). Vždyť také tzv. "živé zkameněliny", tj. druhy, které se dokázaly udržet po desítky až stovky miliónů let naživu, se ukázaly být spíš omylem. S tímto odhalením přišla právě ona relativně mladá makroevolucionistická škola (populární Stephen Jay Gould, Niles Eldredge, Steven Stanley aj.), jejímž příslušníkem se stal i David Raup. "Živé zkameněliny" patří spíš do 40tých až 60tých let tohoto století, do éry "moderní syntézy", kdysi jediného "správného" evolucionistického směru, dnes zdá se definitivně poraženého.
Je ale dnes všechno v pořádku? Zasvěcenec by si snadno mohl všimnout, že Raupově statistice schází některé velmi důležité aspekty. Jak je možné, že rychlost vymírání jednotlivých druhů je uvnitř některých taxonomických skupin zhruba konstantní? Lze to přejít statisticko-sofistickým poukazem na "bankrot hazardního hráče ve hře o život"? Není pohled současné vědy jen pouhým dogmatem, které se zcela iracionálně traduje od dob první mýtické představy o nesmrtelnosti? Ekologové vědí již hodně o tzv. vnitřních faktorech při vymírání druhů, ale evoluční paleontologové se jejich vědomostmi příliš nezatěžují.
Jenže vnitřní zákonitosti existují a velikost populace, jako jedna z nich, není jediným vodítkem. Zajímavé je zhruba podobné průměrné stáří v rámci určitých taxonomických skupin. Je komplikované analogickými čísly v rámci daného typu metabolismu, anebo v rámci dané velikosti těla. Zajímavý je vliv spotřeby kyslíku nejen na věk jedince, ale i na různé trvání druhů ve fosilním záznamu (viz McAlester 1970). Nejsou tato a podobná data dána tím, že nejen v rámci určitého biologického druhu je předurčena zcela konkrétní průměrná délka života jedinců, ale i uvnitř dané fylogenetické linie je analogicky průměrná délka existence druhů tak trochu předurčena, a to bez ohledu na nekonečně proklamované "řídící vlivy prostředí" a oblíbené impakty?
Po přečtení Nulandovy knihy můžeme analogicky tvrdit, že starý člověk také zemře na nemoc, které by jinak snadno odolal, kdyby byl o něco mladší. Mnoho lékařů může po faktické stránce potvrdit, že smrt starého člověka není způsobena nemocí. Nemoc je v podstatě jen jedním z průvodních jevů stárnutí. Je to sice jen úvaha, ale není příliš sofistické tvrdit, že devadesátiletý člověk zemřel na chřipku? Vždyť chřipka nemohla z hlediska konce jeho života způsobit nic jiného, než že ke smrti došlo o něco málo dříve. Aplikujeme-li tuto filosofii na vymírání druhů, můžeme vytvořit teorii vymírání o poznání víc spjatou s fakty než je statistika karetních her" [12].
VÝZNAMNÁ FOSILNÍ NALEZIŠTĚ
Slavné Burgesské břidlice (Burgess Shales) jsou mladší než ediakarská a vendská fauna a spadají již do kambria, tedy prvohor. Jedná se vlastně o naleziště fosilií, které se nachází v Kanadských Skalistých horách v Britské Kolumbii. Je staré 505 milionů let (střední kambrium) a představuje tak jedno z nejstarších nalezišť fosilií živočichů na Zemi. Kromě trilobitů zde byli nalezeni roztodivní členovci, červi, medůzovci a těžko zařaditelní podmořští živočichové, kteří se doposud vymykají nám známým klasifikačním tabulkám. K těm nejzajímavějším patří např. z trilobitů: Bathyuriscus rotundatus, Burlingia hectori, Chancia palliseri, Ehmaniella burgessensis, Elrathia permulta, Elrathina cordillerae, Hanburia gloriosa, Kootenia burgessensis, Ogygopsis klotzi, Olenoides serratus, Oryctocephalus reynoldsi, Pagetia bootes, Ptychagnostus praecurrens. Trilobiti jsou odsud sice velmi zajímaví, ale co činí z burgesských břidlic něco výjimečného, je asociace prapodivných živočichů, nejrůznějších bizarních tvarů, tělní struktury a přívěsků. Tvůrci sci-fi efektů, mimozemských nestvůr a emzáků se nejednou inspirovali právě v této nezvyklé fauně. Jsou to především tito Marrella splendens, Hallucigenia sparsa, Micromitra burgessensis, Leanchoilia superlata nebo Anomalocaris (obr. 14).

Obr. 20 - Zleva: Marrella, Hallucigenia, Micromitra, Leanchoilia a Anomalocaris (rekonstrukce)
Burgesské břidlice objevil americký paleontolog Charles Doolittle Walcott – v roce 1909 odsud popsal první podivné a dosud neznámé druhy kambrických živočichů a postupně vytvořil rozsáhlou sbírku. Zásadní význam tohoto místa rozpoznal později Harry Blackmore Whittington, který také koordinoval další výzkum. Veřejnost se o tomto místě mohla dozvědět např. ze slavné knihy It's a Wonderful Life (1989) autora S. J. Goulda.
Podrobněji jsou burgesské nálezy zpracovány
zde.
Naleziště burgesského typu
K podobným nalezištím označovaným jako „naleziště burgesského typu“ patří Chengjiang v Číně, Kangaroo Island u Austrálie, Sinsk na Sibiři, Buen v Grónsku, ale i část barrandienu v ČR. Jde například o spodnokambrické naleziště v Brdech - souvrství holšinsko-hořické, kde byl objeven záhadný členovec Kodymirus vagans, Kockurus grandis nebo korýš Vladicaris subtilis (Chlupáč 1995) - (obr. 15). Vědecká studie z května 2010 ukázala, že tzv. Burgesská fauna na území dnešního Maroka (souvrství Fezouata - Fezouta) nevyhynula na konci kambria, nýbrž přežila ještě (přinejmenším) do ordoviku. Ukazuje se, že kambrium pozvolna vplulo do ordoviku. Mnoho živočichů se změnilo, mnoho druhů zaniklo, ale mnoho druhů přežilo a metamorfovalo. Podobné nálezy, jako z kambrických burgesských břidlic, jsou známy i z barrandienu, sice ne v takové míře, ale ví se o nich již od dob Barranda z 19 stol. Zajímavé je, že jsou známy z ordovických nálezů, takže tato souvztažnost mezi kambriem se sice předpokládala, ale vědci byli velmi opatrní a v podstatě čekali až na podobný nález, jako je souvrství Fezouta. Mám na mysli problematické ordovické členovce nalézané především v letenském souvrství a to Furca bohemica, Duslia insignis, Xiphosurida sp., Zonozoe drabowiensis, Zonoscutum solum, Caryon bohemicum nebo například Nothozoe barrandei.

Obr. 21 - Ukázka české spodnokambrické fauny. Zleva: Kodymirus vagans, Kockurus grandis (hlavový štít) a Vladicaris subtilis.
Jiř od 19. století byly v křemencích a křemitých pískovcích českého středního ordoviku nalézány zkamenělé zbytky prapodivného tvaru (jednalo se o okolí Berouna například na lokalitách Děd, Veselá, Ostrý). Ze střední plošky nepravidelně čtyřúhelníkového nebo trojúhelníkového tvaru vybíhaly čtyři dlouhé, šavlovitě ohnuté výběžky, končící hrotem. Někdy byly okraje jemně rýhovány, jako by byly roztřepené. Badatelé se dlouho nemohli shodnout na tom, kam záhadné zbytky zařadit, protože nepřipomínaly nic, co by znali nebo alespoň co by něco známého vzdáleně připomínalo. Nakonec je J. Barrande nazval podle jejich vidličnatého tvaru Furca bohemica (furca v latině znamená vidlice). Považoval je sice za zbytky členovců, ale myslel si, že jde o nějaké ocasní přívěsky korýšů. Záhada začala mít jasnější kontury až v roce 1909, kdy již zmíněný Charles Doolittle Walcott objevil v západní Kanadě tzv. burgesské břidlice ze středního kambria s mimořádně zachovanou faunou. Mezi jinými exempláři objevil i záhadného drobného členovce (1,5 - 2 cm) s hlavovým štítem podobně vidličnatého tvaru. Popsal jej pod jménem Marella splendens (obr. 14). Později se ukázalo, že jsou to představitelé specializované větve vodních členovců, vzdálených příbuzných trilobitů. Furca bohemica patří tedy s největší pravděpodobností mezi marrellofoní členovce a její zbytky nejsou ocasní přívěsky, ale hlavové štíty poměrně velkého druhu těchto členovců. Jemnější části krunýřů se v hrubozrných horninách nedochovaly, a tak známe pouze tyto hlavové štíty velké až 5 cm. Jejich zkameněliny jsou známy jen z našeho území (letenské souvrství), i když problematické nálezy se objevují i ve Francii a severní Africe.
Duslia insignis představuje dalšího záhadného členovce ordoviku. Byla popsána již v roce 1903, ale ani autor, ani jiní badatelé si s ní nevěděli rady, a tak ji řadili buďto k pravým korýšům, trilobitům nebo k měkkýšům (chrustnatkám). Problém byl vyřešen v teprve nedávné době, kdy bylo zjištěno, že schránky patří členovcům, podobným trilobitům. Pocházejí ze stejného souvrství jako Furca bohemica, tedy letenského. Tenké, ploché, až 10 cm dlouhé krunýře byly na rozdíl od krunýřů trilobitů z chitinofosfatického materiálu (podobně jako u konulárií) a nebyly schopny stáčení (volvace). Mají široce eliptický tvar s dobře odlišitelným hlavovým štítem, článkovaným tělem a drobným, plutvičkovitým ocasním štítem (telsonem). Střední osa těla byla méně výrazná než u trilobitů. Kolem okraje těla je vyvinut třásnitý lem. Ten nejspíš pomáhal Duslii při pohybu. Otisky a jádra jejich krunýřů se v hornině nacházejí vždy celé v tzv. životní pozici, pravděpodobně břišní stranou dolů [13].
Obr. 22 - Ordovická problematika českého barrandienu. Zleva: Furca bohemica (otisk a rekonstrukce hlavového štítu), Duslia insignis (otisk a rekonstrukce kompletního členovce), Zonozoe drabowiensis (rekonstrukce hlavového štítu).
MALÁ SPEKULACE NA ZÁVĚR - věk trilobitů
Jak je to vlastně s předpokládanou délkou života trilobitů? To je jistě otázka, kterou si položil nejeden paleontolog. Odpovědět na ni jednoznačně však nelze, už jen pro to, že trilobiti měli několik vývojových stádií, které mohli mít u různých druhů, různou časovou délku. Jsou to tato stadia (mírně zjednodušeno) - protaspidní (larvální) stadium, meraspidní stadium (první trupové články), holaspidní stadium (dospělý jedinec) - podrobněji o tom výše, v části ONTOGENEZE - růst a vývoj. Je možné, že některé druhy prožily v larválním stádiu většinu života, aby v dospělém rychle zplodili potomky a uhynuli, tak jak to známe u dnešních jepic. A to je právě ono, jediná věc o kterou se můžeme opřít je věk recentních členovců, tedy nepřímých potomků této velké vyhynulé podtřídy mořských živočichů. Věk u recentních členovců je velmi různý, od několika hodin (jepice /Ephemeroptera/ v dospělém stadiu, v larválním cca 1-3 roky, podle druhu) až po mnoho desítek let (humr /Homarus/). Například o věku současných krabů není moc dostupných informací. Jsou druhy, které umírají po jednom roce a naopak jsou druhy, které v pěti letech dosáhnou pohlavní zralosti. U jednoho druhu lze zralost ovlivnit i teplotou vody. Různá teplota přináší i různou dobu vyspělosti. Druh suchozemského kraba Cardisoma armatum se dožívá v přírodě okolo 10-15let. Naproti tomu mořští humři se dožívají mnoha desítek let. Doložený případ velmi vysokého stáří humra je 140 let. Rekordmany v dlouhověkosti jsou mezi bezobratlými také sklípkani /Mygalomorphae/ – dožívají se 25 až 30 let. Takto dlouho žijí ale pouze samičky. Sameček má před sebou od okamžiku, kdy dospěje (což bývá tak ve dvou až třech letech), už jen rok, rok a půl. Jeho úkolem je najít samičku a oplodnit ji. Poté většinou po několika dnech až týdnech umírá, někdy jeho život ukončí samotná samička. Velmi zajímavá je i délka života recentních ostnokožců, např. běžné ježovky Strongylocentrotus franciscanus ze západního pobřeží severní Ameriky. S nedávným poznatkem o věku těchto ježovek přišli američtí vědci z Oregon State University spolu s kolegy z Lawrence Livermore National Laboratory. Pohlavní dospělosti dosahují sice již do druhého roku života, poté se však jejich stárnutí zpomalí, až téměř zastaví. Podle odhadů založených na radiokarbonové metodě se mohu některé exempláře, žijící v příhodných podmínkách dožívat až stáří 200 let.
Jak je to tedy s délkou života u trilobitů? Můžeme li vycházet ze spekulativních poznatků a našich vědomostí týkajících se délky věku recentních členovců, mohli se někteří dožívat jednoho dne, jiní zajisté mnoho desetiletí. Je známo přes 15 000 druhů trilobitů (některé zdroje uvádějí přes 17 000), což je obrovská diference ontogenetická, ve způsobu života i morfologická a navíc toto číslo neustále narůstá. Takže délka života se opravdu mohla pohybovat od "mikrověku" k "makrověku", druh od druhu. Ještě jedna malá spekulace na závěr. Mlž Panopea generosa ze západního pobřeží Severní Ameriky se může dožít až 168 let nebo jiný mlž druhu Mercenaria mercenaria dokonce 405 let. U mušlí se věk pozná poměrně snadno – mají totiž podobně jako stromy letokruhy. Co když ale takové letokruhy měli i trilobiti. Namátkou si vybavuji hned jednoho, známého ze šáreckého souvrství. Myslím trilobita Ectillaenus katzeri (Barrande, 1872)†. Na hlavě i pygidiu tohoto trilobita, je v příznivém zachování rozeznatelná struktura, podobná gramofonové stopě, připomínající letokruhy. Myšlenka svádí k tomu, zdali to není možný záznam po růstu trupu trilobita a zdali by se tak jeho věk "jednoduše" nedal spočítat.

Obr. 23 - detail hlavového (vlevo) a ocasního štítu trilobita Ectillaenus advena s možnými stopami po růstu. Zdroj: https://ordovik.sweb.cz/
APENDIX
Trilobiti jsou známi výhradně z prvohorních mořských sedimentů. Největšího rozkvětu dosahují již ve středním kambriu a v ordoviku. Po mírném ústupu v období siluru následuje další rozvoj v devonu, kdy se objevuje i řada zcela nových typů. Do poslední periody prvohor - permu, přežívá jen jedna jediná čeleď Phillipsiidae. S koncem permu však mizí i tito poslední zástupci. Do současné doby bylo popsáno více než 15 000 druhů trilobitů z celého světa, z toho přes 1500 z Čech!
V Čechách nacházíme trilobity především v klasickém území Barrandienu (území rozprostírající se od Kralup, přes Kladno, Stříbro, Klatovy, Příbram, Říčany až po Brandýs nad Labem), ale vyskytují se i v paleozoiku - prvohorách Železných hor, Jeseníků a Drahanské vrchoviny, v devonu Moravského krasu a na velmi významné lokalitě Čelechovice na Hané. Jen v Barrandienu je to okolo 1500 druhů. Na Slovensku lze nalézt trilobity v karbonských břidlicích u Ochtiné, Dobšiné a Jelšavy v slovenském Rudohoří (např. trilobita Weania rozlozsniki RAKUSZ, Paladin eichwaldi (FISCHER), Phillipsia margaritifera ROEM, Cummingella aff. Balladoolensis Red, Cyrtoproetus dobsinensis Illés nebo arachnida Anthracomartus voelkelianus Karsch (Hyžný et al. 2013), dále nautiloidy, krionidy, přesličky, kapradiny atp.
Zdroje:
Rudolf Prokop. Zkamenělý svět, vydala Práce v Praze, ISBN 80-208-0888-4, 1989 [1], [4], [5], [13]
https://www.wikipedia.org [2], [3]
Lukáš Laibl. Ontogeneze trilobitů, - bakalářská práce, Přírodovědecká fakulta Univerzity Karlovy v Praze [6], [7], [8], [9], [10], [11]
Václav Petr. JAK DRUHY VYMÍRAJÍ, publikováno v časopise Český kras, 23: 54-57. Beroun 1997 [12]





